Zenith is a first-of-its-kind next-generation “proto-biotic” supplement that unites probiotics, prebiotics, postbiotics, and phytobiotics into one comprehensive formula to optimize the entire gut-brain axis. Backed by clinical evidence and patented botanical synergies, zenith helps reduce stress and anxiety, sharpen cognition, improve circadian sleep/wake cycles, and fortify metabolic health*. It’s not just about hitting peak performance once – it’s about showing up with zen, resilience and everyday excellence.
Zenith is a next-generation “multi-biotic” dietary supplement designed to optimize the microbiome–gut–brain axis (MGBA) and deliver daily excellence in stress resilience, mood balance, cognitive clarity, restorative sleep, and metabolic vitality. Unlike conventional probiotics, zenith combines four complementary categories of “biotics”—probiotics, prebiotics, postbiotics, and phytobiotics—to create a synergistic ecosystem approach.
Probiotic Blend (BifiZen® – 4 Billion CFU)
Prebiotic Blend (PhytoSpherix® & MicrobiomeX®)
Phytobiotic Blend (Vigortonin®, Enzogenol®, Venetron®)
Postbiotic Blend (Suntheanine® & Akkermansia muciniphila AKK001)
Zenith is a next-generation dietary supplement (“Proto-Biotic”) that integrates four complementary “biotic” blends — probiotic, prebiotic, postbiotic, and phytobiotic – designed to harmonize the entire Gut-Brain-Axis, support stress resilience, and optimize mental and physical performance.
Zenith represents a scientifically grounded, multi-biotic strategy that targets the microbiome–gut–brain axis with complementary approaches: prebiotic fueling, phytobiotic signaling, postbiotic neuro-support, and probiotic colonization. Together, these ingredients form a synergistic ecosystem to enhance daily excellence – supporting stress resilience, mood balance, cognitive clarity, and restorative sleep.
This white paper summarizes the mechanistic rationale, clinical evidence, and key benefits of each branded research-supported ingredient in the zenith formula.
Emerging scientific evidence demonstrates that specific strains of probiotics can modulate the gut-brain axis and meaningfully influence psychological well-being, sleep quality, and cognitive-emotional resilience.
The gut microbiome communicates bidirectionally with the central nervous system via the gut-brain axis, integrating neural, immune, and endocrine signaling pathways. Certain probiotics can modulate neurotransmitter synthesis (e.g., GABA, serotonin), stress hormone regulation (cortisol), and inflammatory pathways, all of which impact mood, sleep, and stress responses. Clinical evidence has moved beyond theory, now providing quantifiable effects on sleep metrics, mood indicators, and stress reactivity.
The 4-strain Bifizen™ blend—Limosilactobacillus fermentum LF16, Lacticaseibacillus rhamnosus LR06, Lactiplantibacillus plantarum LP01, and Bifidobacterium longum BL04 has shown a range of mental wellness benefits in healthy adults (4×10⁹ CFU/day for 6 weeks).
Across two randomized, double-blind, placebo-controlled trials in healthy adults, this exact 4-strain blend (1 billion CFU of each strain) improved validated measures of mood and anxiety and boosted circulating serotonin, as well as improved subjective sleep quality (e.g. falling asleep faster and fewer disturbances).
For generally healthy, stressed-but-functioning adults, this 4-strain “psychobiotic” pattern looks like calmer mood, less anxious tone, and smoother bedtime—with a mechanistic fingerprint of higher circulating serotonin and subjective sleep gains.
Limosilactobacillus fermentum LF16
Lacticaseibacillus rhamnosus LR06
Lactiplantibacillus plantarum LP01
Bifidobacterium longum BL04
Marotta et al. (2019) – Cognitive Reactivity, Mood, and Sleep
Irwin et al. (2020) – Systematic Review and Meta-Analysis of Probiotics on Sleep
Calgaro et al. (2021) – Microbiota Modulation with Probiotics
Walden et al. (2023) – Probiotics and Mood Biomarkers
Across diverse study designs—RCTs, meta-analyses, microbiome sequencing—the evidence converges on four central conclusions:
Mechanistic Pathways
Specific strains of probiotics represent a safe, evidence-based intervention for enhancing psychological resilience, sleep health, and mood balance. When formulated as multi-strain blends with clinically validated strains, probiotics can be positioned as part of an integrated approach to stress management and mental wellness.
The cumulative evidence supports their use in:
CLINICAL STUDIES
1) Marotta A, et al. Effects of Probiotics on Cognitive Reactivity, Mood, and Sleep Quality. Front Psychiatry. 2019 Mar 27;10:164. doi: 10.3389/fpsyt.2019.00164.
2) Irwin C, et al. Effects of probiotics and paraprobiotics on subjective and objective sleep metrics: a systematic review and meta-analysis. Eur J Clin Nutr. 2020 Nov;74(11):1536-1549. https://doi.org/10.1038/s41430-020-0656-x
3) Calgaro M, et al. Metabarcoding analysis of gut microbiota of healthy individuals reveals impact of probiotic and maltodextrin consumption. Benef Microbes. 2021 Apr 12;12(2):121-136. doi: 10.3920/BM2020.0137
4) Walden KE, et al. A randomized controlled trial to examine the impact of a multi-strain probiotic on self-reported indicators of depression, anxiety, mood, and associated biomarkers. Front Nutr. 2023 Aug 31;10:1219313. doi: 10.3389/fnut.2023.1219313
5) Kerksick CM, et al. (2024). Multi-strain probiotic improves subjective sleep quality with no impact on body composition, hemodynamics, and physical activity. Beneficial Microbes. DOI: 10.1163/18762891-bja00002 (NCT05343533). https://dx.doi.org/10.1163/18762891-bja00002
PhytoSpherix® (Phyto-glycogen, Zea mays leaf extract)
• Mechanism: Plant-based glycogen acts as a fermentable carbohydrate, fueling beneficial microbiota and supporting SCFA (short-chain fatty acid) production.
• Evidence: Preclinical studies demonstrate glycogen fermentation into acetate, propionate, and butyrate, with downstream impacts on energy metabolism and neurotransmitter precursors (serotonin, GABA)【Selleck, 2020】. Early human safety/tolerance data confirm its GRAS status.
• Benefits: Supports microbiome resilience, metabolic flexibility, and energy balance.
Substrates that are selectively used by host microorganisms of the gut microbiota, which have the potential to improve human and animal health, are considered prebiotics (1). In vitro simulations can provide detailed insights into the effects of test ingredients on gut microbial composition and functioning. One such simulation is ProDigest’s Colon-on-a-Plate® (CoaP), an advanced in vitro system that simulates human intestinal microbial activity, enabling the study ofthe effects of drugs, probiotics, and other compounds on gut microbiota (2).
The gut microbiome is known to produce small molecules, so-called metabolites, that impact human function. One of these metabolite groups include the short chain fatty acids (SCFAs), which are produced by bacterial fermentation of non-digestible polysaccharides in the large intestine (3). Extensive evidence indicates that SCFA signaling can affect the gut and the central nervous system (CNS) (3-5). In the gut, the SCFA can improve intestinal health, including the maintenance of intestinal barrier integrity and protection against inflammation (6, 7). Regarding the gut-brain axis, the SCFAs can affect the CNS by modulating the levels of neurotransmitters and regulating the expression of tryptophan-5-hydroxylase, an enzyme which is involved in serotonin synthesis (4, 8, 9). Serotonin is a neurotransmitter that plays a crucial role in mood regulation, along with other functions such as sleep, appetite, and digestion (10). Furthermore, SCFA are absorbed via monocarboxylate transporters (MCTs) and the abundant expression of MCTs on brain endothelial cells suggest that they can cross the BBB. This is further supported by the presence of SCFAs in human cerebrospinal fluid (11). There is growing evidence that SCFAs may modulate the hypothalamic-pituitary-adrenal (HPA) axis, which is the body’s main stress response system (12) (Figure 1).
Figure 1: Effects of Dietary Fiber on Gut-Brain Axis
Intriguingly, the complex and dynamic human metabolome, a compilation of all primary and secondary metabolites, can be influenced by factors like microbial community and dietary patterns (13). High-resolution mass spectrometry techniques are used for metabolic fingerprinting and can give in-depth information on a metabolic system. As such, metabolomics can provide a comprehensive analysis of metabolites, yielding a direct functional read-out of the physiological status of an organism and the possible impact of a dietary intervention, such as PhytoSpherix supplementation (14). In this context, with the microbiological community being an important factor, insights on how to modulate the composition and/or metabolic activities of the microbiome are valuable to support a metabolically healthy condition (Figure 1).
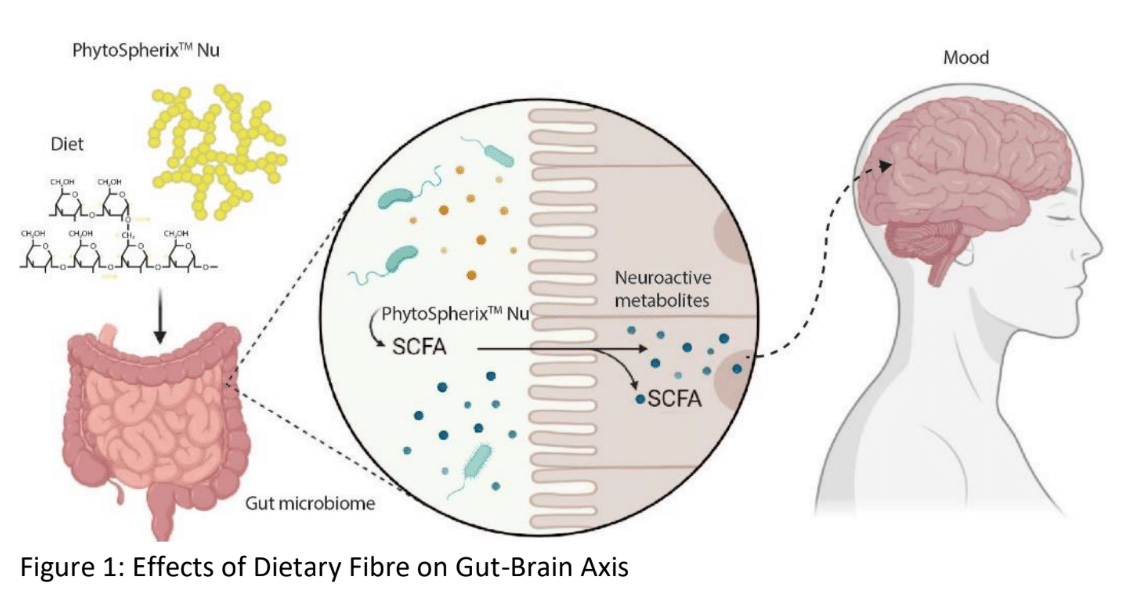
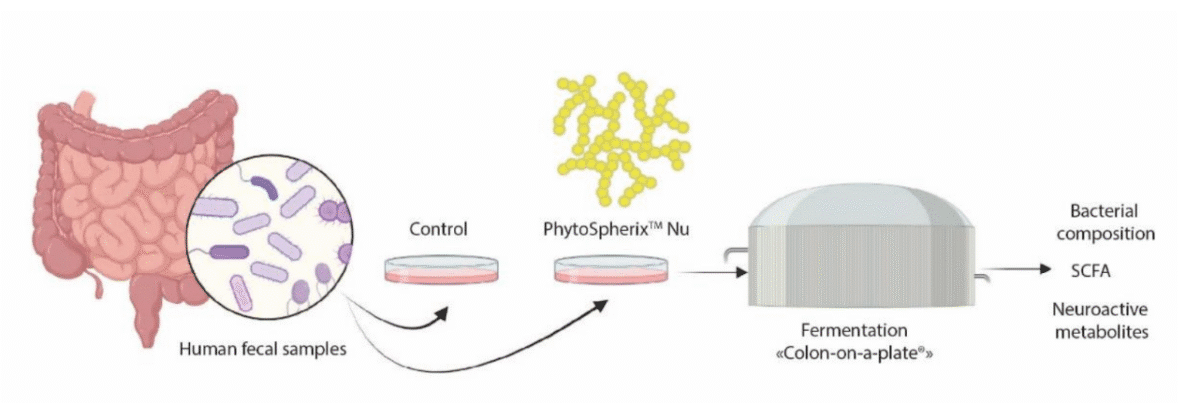
Figure 2: Experimental design of Colon-on-a-Plate® simulation
PhytoSpherix is a natural phytoglycogen derived from a special variety of sweet corn that has been evaluated for its effects on the colonic microbiome healthy subjects using ProDigest’s Colon-on-a-Plate® simulation platform, where the human colon was mimicked with an intricate system ex vivo. The effect of PhytoSpherix on the microbial metabolic activity was determined by targeting saccharolytic markers such as short-chain fatty acids (SCFA) and lactate, as well as neuroactive compounds. It’s impact on microbial community composition was assessed in human fecal samples (n=10) with 16S targeted Illumina sequencing.
From the experimental model it was evident, that PhytoSpherix has positive effects on the gut microbiome. The phytoglycogen was efficiently fermented by the gut microbiota and a significant increase in SCFA including acetate (+94 %, p<0.0001), propionate (+40 %, p<0.01) and butyrate (+96 %, p=0.0516) was observed, demonstrating the prebiotic efficiency of PhytoSpherix (Figure 3).
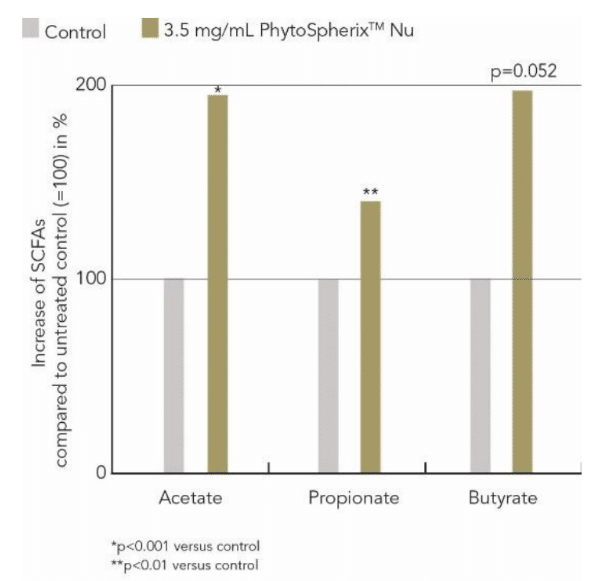
Figure 3: Stimulation of SCFAs by PhytoSpherix
PhytoSpherix treatment significantly stimulated bacterial biomass production, and this was consistently observed for each individual donor, including significant enrichments of health-beneficial bacteria including Bifidobacterium, Faecalibacterium (Ruminococcaceae), and members of the Erysipelotrichaceae and Lachnospiraceae families. Again, the effect was observed and consistent across all individual donors. In addition, the analysis revealed increased abundances of Streptococcus and Enterococcus and members of the Veillonellaceae family.
When evaluating the data in more detail, a positive association was found between stimulation of the SCFA propionate and the enrichment of Veillonellaceae, Lactobacillaceae, Ruminococcaceae, Coriobacteriaceae and Erysipelotrichaceae. Interestingly, Veillonellaceae contains bacterial species able to produce propionate from lactate, while Lactobacillaceae, Coriobacteriaceae and Erysipelotrichaceae produce lactate (which in turn can be converted to propionate). Production of acetate was strongly correlated with Bifidobacteriaceae, and butyrate-production was strongly linked with presence of Enterococcaceae and Streptococcaceae.
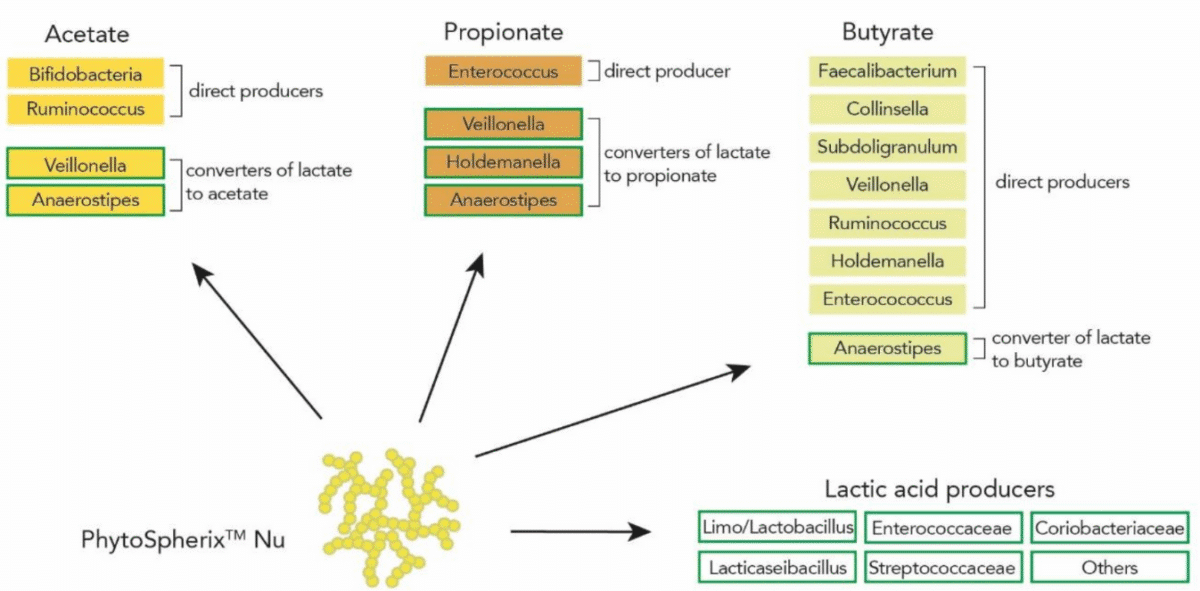
Figure 4: Enrichment of bacteria after PhytoSpherix treatment
When evaluating the neuroactive metabolites, it was observed that PhytoSpherix affected 12 neurotransmitters, which play a role in the gut-brain (and body) axis
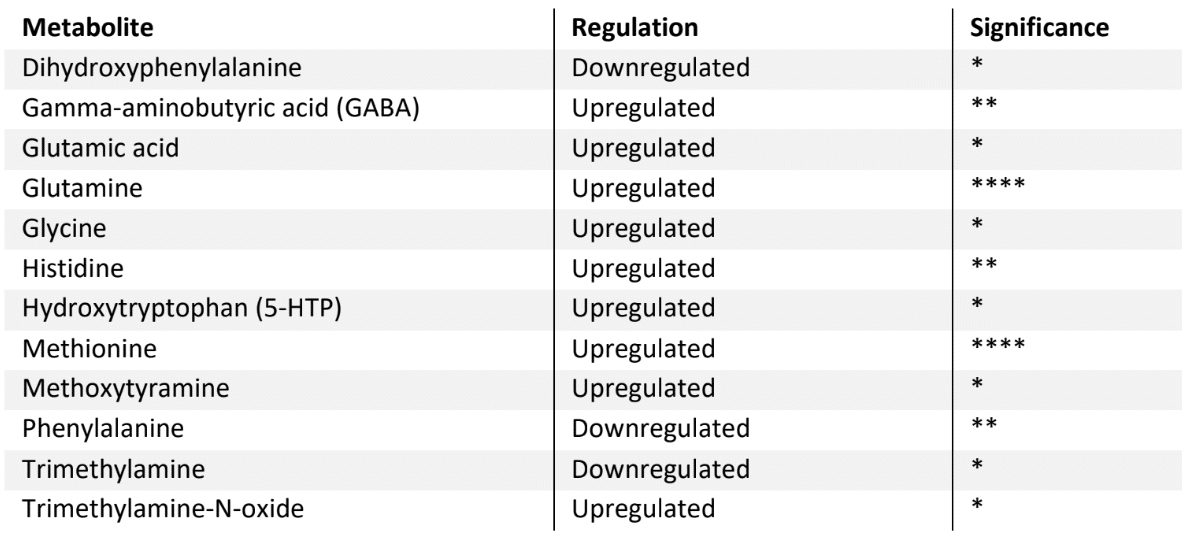
Table 2: Neuroactive metabolites affected by PhytoSpherix treatment
Compared to untreated controls, the treatment with PhytoSpherix significantly upregulated gamma-aminobutyric acid (GABA). Increased GABA levels can reduce anxiety, promote relaxation, and improve overall mood (16). Higher GABA levels are associated with better sleep quality. GABA helps to quiet the mind and prepare the body for sleep, reducing insomnia and improving sleep patterns. Indeed, some research indicates that GABA produced by gut bacteria may influence brain function and mood (17).
Further, the amino acid Glutamine was significantly stimulated with PhytoSpherix treatment. Glutamine is a precursor to glutamate, an excitatory neurotransmitter, and GABA, an inhibitory neurotransmitter. Both are critical for brain function and mood regulation ref, as such upregulating glutamine could potentially improve mood and reduce symptoms of anxiety and depression. Increased glutamine levels can also support cognitive functions such as learning and memory by maintaining a balance between excitatory and inhibitory neurotransmitters (18).
PhytoSpherix further stimulated glycine, a non-essential amino acid that serves as a neurotransmitter in the central nervous system and is known to improve sleep quality (19).
Further stimulated neuroactive metabolites include methionine, which is involved in the synthesis of SAMe (S-adenosylmethionine, an important regulator of mood and cognitive function), and methoxytyramine, which is a metabolite of dopamine (20).
Importantly, hydroxytryptophan (5-HTP) was significantly upregulated after PhytoSpherix treatment. 5-HTP is a precursor to serotonin, an important neurotransmitter regulating mood, anxiety and sleep – and is often used to help manage depression and anxiety. By increasing serotonin levels, 5-HTP can also boost melatonin production, thereby improving sleep quality (21).
In summary, this evidence positions the natural and safe compound as an ideal substrate to improve the mood via the gut-brain axis.
MicrobiomeX® (Flavobiotics from Citrus sinensis and Citrus paradisi)
Citrus flavonoid extracts help reshape microbial communities, particularly boosting acetate-producers and butyrate levels, quell inflammatory signaling, and tilt microbial metabolism toward beneficial SCFA production.
Title: Sost et al. A Citrus Fruit Extract High in Polyphenols Beneficially Modulates the Gut Microbiota of Healthy Human Volunteers in a Validated In Vitro Model of the Colon. Nutrients, 2021; 13(11):3915. https://doi.org/10.3390/nu13113915
Summary: Using the TIM‑2 dynamic in vitro colon model, the researchers investigated the impact of a citrus fruit extract rich in hesperidin and naringin (250–350 mg/day for 3 days) on gut microbial composition and activity. They observed a dose-dependent increase in beneficial bacteria such as Roseburia, Eubacterium ramulus, and Bacteroides eggerthii, along with elevated acetate production. Increased distal-gut acetate may boost fat oxidation and satiety—a promising mechanistic hint for metabolic health.
Title: Stevens et al. The effects of citrus flavonoids and their metabolites on immune-mediated intestinal barrier disruption using an in vitro co-culture model. British Journal of Nutrition, 2022;128(10):1917–1926. https://doi.org/10.1017/s0007114521004797
Summary: Leveraging a Caco-2/THP-1 immune co-culture model mimicking immune-mediated gut barrier stress, this study tested citrus flavonoids (like hesperidin/naringin) and their metabolites. There was significant inhibition of NF‑κB activity and pro-inflammatory cytokine release (IL‑8, TNF‑α, IL‑6)—though IL‑1β oddly increased.
Title: Sost et al. Citrus Extract High in Flavonoids Beneficially Alters Intestinal Metabolic Responses in Subjects with Features of Metabolic Syndrome. Foods, 2023; 12(18):3413. https://doi.org/10.3390/foods12183413
Summary: This combined in vitro/in vivo study tested a flavonoid-rich citrus extract (500 mg/day) in the TIM‑2 model and in human subjects with metabolic syndrome features. In both settings, short-chain fatty acid (SCFA) production (butyrate, acetate, valerate) significantly increased. In human participants over 12 weeks, there was a notable rise in butyrate levels (p = 0.022) and a trend toward lower fecal calprotectin (a gut inflammation marker, p = 0.058) . These metabolic shifts hint at improved gut—and possibly systemic—health.
Study & Model | Intervention | Key Findings |
Sost et al. 2021 (in vitro) | Citrus polyphenol extract (250–350 mg/day) | ↑ Roseburia, E. ramulus, B. eggerthii; ↑ acetate |
Stevens et al. 2022 (in vitro) | Citrus flavonoids & metabolites | ↓ NF-κB, ↓ IL-8/TNF-α/IL-6, ↑ IL-1β; no barrier integrity change |
Sost et al. 2023 (in vitro + in vivo) | Citrus flavonoid extract (500 mg/day) | ↑ SCFAs (butyrate, acetate, valerate); in humans: ↑ butyrate, ↓ fecal calprotectin (trend) |
The health benefits of MicrobiomeX® have been assessed in two randomized double-blind clinical studies, which demonstrated its beneficial effect on the gut and immune health. Daily consumption of 500mg of MicrobiomeX® beneficially modulates the gut microbiota. The growth of specific bacterial groups, namely Clostridium cluster XIVa is promoted and particularly the species belonging to the genus Roseburia spp, which are well-known for their butyrate-producing property and many beneficial health effects.
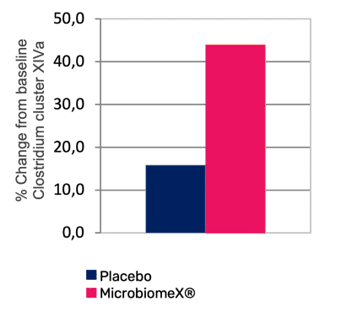
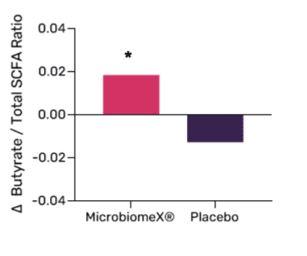
Short-Chain Fatty Acid Profile
After 12 weeks of 500mg MicrobiomeX® consumption, a significant increase in the butyrate proportion of the total SCFA was observed. Furthermore, the butyrate over acetate ratio significantly increased in the MicrobiomeX® group compared to the placebo group. Butyrate has health-promoting effects such as positively affecting gut barrier function and lowering gut inflammation.
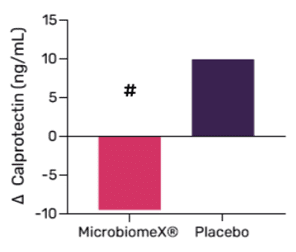
Gut Inflammation & Immune Health
After 12 weeks of daily supplementation with 500mg of MicrobiomeX®, a strong reduction in faecal calprotectin was shown compared to the placebo group. Calprotectin is an important biomarker of gut inflammation, and its reduction suggests a decrease in pathogen load in the gut.
Vigortonin (Ephedra viridis stem extract)
The FDA’s 2004 Final Rule declared dietary supplements containing ephedrine alkaloids adulterated due to unreasonable safety risks. Crucially, this restriction does not extend to Ephedra species—like E. viridis—that lack those alkaloids. In other words, while Ephedra sinica‑derived products are banned (Chinese ephedra), teas and other products derived from non‑ephedrine species remain legal, provided they do not contain ephedrine alkaloids.
New Zealand pine bark (Pinus radiata) is an adaptogen-like aid for stress resilience, cognitive performance, and glycemic control – behaving as stress-response normalizers that can positively tune the interconnected redox, vascular, and neuroimmune signals that underpin “calm energy” and mental clarity. Due to it being rich in oligomeric proanthocyanidins (OPCs), catechins, and taxifolin – which support vascular function, neuroprotection, and antioxidant defense – benefits include cognitive enhancement, stress buffering, vascular and antioxidant support.
Neurovascular support: OPCs enhance endothelium-dependent vasodilation (↑ nitric-oxide bioactivity), improving perfusion that supports attention and working memory; a human study showed ~41% higher acetylcholine-induced forearm blood flow after 2 weeks (180mg/day).
Redox/immune modulation: Pine bark lowers oxidative stress and inflammatory signaling – mechanisms repeatedly linked to lower perceived stress/anxiety and better cognitive scores in human trials and integrative reviews.
Glycemic & endothelial balance: In people with cardiometabolic risk or type-2 diabetes, pine bark has reduced fasting glucose and HbA1c, alongside improvements in lipids and blood pressure in meta-analysis of RCTs.
Enzogenol® RCT (5 weeks, men 50–65): Faster spatial working memory and immediate recognition vs. control.
Broader Pycnogenol® (Maritime pine, Pinus pinaster) evidence: Reviews summarize RCTs reporting improvements in attention, alertness, episodic and working memory, often alongside drops in oxidative stress markers.
Integrative clinical reviews note reduced anxiety and improved mental performance in healthy adults taking Pycnogenol, consistent with an anti-stress/normalizing profile rather than stimulation. (Mechanism: antioxidant + endothelial + neuroimmune effects).
Meta-analysis of 24 RCTs (n≈1,594): Pycnogenol reduced fasting glucose and HbA1c. 12-week Pycnogenol (100 mg/day) lowered plasma glucose and improved endothelial measures; HbA1c reductions were evident within 1 month in responders and favorable effects on fasting glucose and adhesion molecules over 8 weeks in metabolic risk profiles.
Pine bark is less “caffeine hit,” more circuit-tuner: it opens vessels, cools oxidative noise, and steadies metabolism. The result is an adaptogen-like lift in calm focus, stress resilience, and metabolic housekeeping.
1. Panossian, A. G. (2020). Evolution of the adaptogenic concept from traditional use to modern understanding: an operational definition of adaptogens. [Review Article]. Pharmaceuticals. Available at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7756641/
2. Simpson, T., Kure, C., & Stough, C. (2019). Assessing the efficacy and mechanisms of Pycnogenol® on cognitive aging from in vitro, animal and human studies. Frontiers in Pharmacology, 10:694. Available at: https://www.frontiersin.org/journals/pharmacology/articles/10.3389/fphar.2019.00694/full
3. Pipingas, A., et al. (2008). Improved cognitive performance after dietary supplementation with Enzogenol® and vitamin C: randomized controlled trial. Journal of Psychopharmacology. Available at: https://pubmed.ncbi.nlm.nih.gov/18683195/
4. Weichmann, F., & Rohdewald, P. (2024). Pycnogenol® French maritime pine bark extract in randomized, double-blind, placebo-controlled human clinical studies: overview of effects on endothelial function, diabetes, and other health domains. Frontiers in Nutrition, 11:1389374. Available at: https://pubmed.ncbi.nlm.nih.gov/38757130/
5. Malekahmadi, M., et al. (2019). Effects of Pycnogenol® on cardiometabolic health: systematic review and meta-analysis of randomized controlled trials—fasting glucose and HbA1c outcomes included. Nutrition Research. Available at: https://pubmed.ncbi.nlm.nih.gov/31585179/
6. Liu, X., Wei, J., Tan, F., Zhou, S., Würthwein, G., & Rohdewald, P. (2004). Antidiabetic effect of Pycnogenol® French maritime pine bark extract in patients with type II diabetes: glucose reduction and improved endothelial function. Life Sciences, 74(7):855–862. Available at: https://pubmed.ncbi.nlm.nih.gov/15363656/
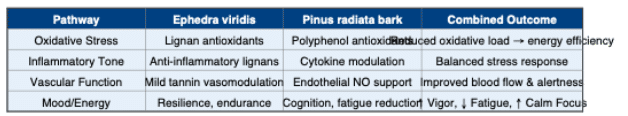
Ephedra viridis (Green Mormon tea or “American ephedra”) and Pinus radiata bark extract represent a unique ephedrine-free adaptogenic combination. While E. viridis provides lignan- and polyphenol-driven resilience without stimulant effects, P. radiata bark contributes vascular, antioxidant, and anti-stress mechanisms.
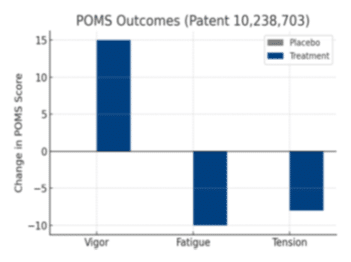
Importantly, U.S. Patent 10,238,703 demonstrates that this blend improves Profile of Mood States (POMS) outcomes, with increased vigor and reduced both fatigue and tension in humans. Together, these botanicals provide calm, sustainable energy and mood balance.
Adaptogen Framework
Adaptogens are defined as natural compounds that normalize physiological function, improve mental and physical performance under stress, and enhance non-specific resistance to stressors. E. viridis and P. radiata align with this definition through redox balance, inflammatory modulation, vascular support, and stress-axis effects without overstimulation.
Ephedra viridis: Stimulant-Free Adaptogen
Ephedra viridis has long been used traditionally as “Mormon tea”; “Brigham tea” and “American” ephedra. Modern analyses confirm that E. viridis lacks ephedrine alkaloids, and that its bioactivity is instead driven by non-alkaloid constituents – such as lignans, tannins, and proanthocyanidins that have well-known antioxidant, anti-inflammatory, and vasomodulatory properties. These mechanisms support mental endurance, fatigue resistance, and resilience under stress, aligning with the adaptogenic model without sympathomimetic risks common to stimulant-based ingredients like caffeine and “Chinese” ephedra (Ephedra sinensis).
Pinus radiata Bark Extract: Cognitive & Vascular Support
Pinus radiata bark extract is rich in proanthocyanidins and catechins, structurally related to Pycnogenol (Pinus pinaster). These compounds support vascular health, endothelial nitric oxide activity, neurovascular coupling, and antioxidant defenses. Clinical studies with pine bark polyphenols show benefits for cognition, stress perception, and fatigue reduction, providing strong rationale for synergy with E. viridis.
Synergy: Ephedra viridis + Pinus radiata (Patent 10,238,703)
The patented combination of E. viridis extract with P. radiata bark extract demonstrated measurable improvements in mood and vigor. Human participants reported significant improvements in Profile of Mood States (POMS) scores, including reduced fatigue and tension and increased vigor. This indicates a synergistic adaptogenic effect: E. viridis modulates oxidative and inflammatory resilience, while P. radiata enhances circulation and cognitive performance. Together, they deliver calm, sustained energy and improved mood state.
Venetron® (Apocynum venetum, Rafuma leaf extract)
Venetron is a plant-based adaptogen extract rich in hyperoside and isoquercitrin that modulates GABAergic and monoaminergic pathways, with no impact on common drug metabolism pathways (CYP3A, P-glycoprotein), and delivering numerous benefits:
Clinical Evidence
Suntheanine® (L-theanine)
Suntheanine® is a clinically validated form of L-theanine that:
Together, these effects make Suntheanine a dual-action adaptogen-like compound—providing relaxation without drowsiness and alertness without overstimulation.
Stress & Cortisol Regulation:
Suntheanine® modulates the HPA axis and neurotransmitters (glutamate, GABA, dopamine, serotonin), lowering cortisol during stress exposure.
Alpha Brain Wave Enhancement:
Multiple studies show L-theanine increases alpha brain wave activity, associated with relaxed alertness, calm focus, and reduced anxiety.
Cognition & Relaxed Alertness:
By reducing stress reactivity while increasing calm focus, Suntheanine supports dual effects of relaxation and mental performance:
Sleep Quality (Indirect):
While not sedative, theanine reduces nighttime stress arousal and improves sleep satisfaction and efficiency, especially in anxious or ADHD populations.
References
Akkermansia muciniphila AKK001 (pasteurized)
AKK001 is a specific, pasteurized (heat-treated) strain of the beneficial gut bacterium Akkermansia muciniphila. It is used as a postbiotic ingredient in dietary supplements aimed at supporting metabolic and gut health. Akkermansia is a “keystone” mucin-degrading bacterium that supports overall gut integrity – and the pasteurized form stimulates immune and metabolic pathways more strongly than live cells. One study of overweight adults (Depommier et al. 2019) found that pasteurized Akkermansia improved insulin sensitivity, reduced plasma cholesterol, and enhanced gut barrier integrity.
Pasteurized A. muciniphila (AKK001) is best viewed as a barrier-first, immune-calibrating postbiotic with human-level metabolic benefits (↑ insulin sensitivity; ↓ total cholesterol) and strong mechanism for impacting weight/GLP-1 physiology.
Because AKK001 is pasteurized, it contains no live bacteria. This makes it a “postbiotic” – meaning it delivers the beneficial compounds produced by the bacteria without the need for colonization, unlike traditional probiotics. AKK001’s unique properties may offer different or more pronounced benefits than other Akkermansia strains.
AKK001 leverages the well-researched benefits of Akkermansia muciniphila, which is associated with:
• Improved Metabolic Health and Weight Management: Research suggests a link between healthy levels of Akkermansia and better glucose metabolism, insulin sensitivity, and reduced body fat.
• Reinforced Gut Barrier: This strain’s bioactive outer membrane proteins are thought to strengthen the gut lining, which is crucial for digestive health and a strong immune response. Barrier integrity → lower endotoxemia & inflammatory tone → improved insulin sensitivity.
• Reduced Inflammation: Akkermansia is known to reduce inflammation within the gut microbiome.
• Immune homeostasis (IL-10/TLR2; commensal antigen education) → calmer mucosal milieu supportive of metabolic efficiency.
• Enteroendocrine (GLP-1) & thermogenesis (preclinical) → appetite/glycemic & energy-expenditure effects that complement barrier and immune mechanisms.
Pasteurized A. muciniphila has emerged as a next-generation, barrier-centric postbiotic with human evidence for improving insulin sensitivity and lowering insulinemia and total cholesterol in insulin-resistant adults—without changing diet or activity. Mechanistically, it fortifies the mucus layer and tight junctions, normalizes inflammatory tone, and engages enteroendocrine signaling (GLP-1) in preclinical models, helping explain downstream effects on glycemic control, metabolic efficiency, and weight regulation. Safety has been vetted by EU regulators for adult use at defined cell equivalents.
A. muciniphila is a mucin-specialist that resides at the intestinal mucus interface. Pasteurization preserves surface structures (e.g., Amuc_1100) that signal through TLR2 and related pathways; in mice, pasteurized cells outperform live cells for improving adiposity, insulin resistance, and dyslipidemia, pointing to cell-surface–mediated postbiotic activity.
Randomized, double-blind, placebo-controlled proof-of-concept (3 months; overweight/obese, insulin-resistant adults; 10¹⁰ cells/day):
• ↑ Insulin sensitivity (~+29% vs placebo) and ↓ fasting insulin (~−34% vs placebo).
• ↓ Total cholesterol (~−8.7% vs placebo).
• Trends toward lower body weight and fat mass.
• In diet-induced obesity, A. muciniphila reverses metabolic endotoxemia, increases mucus thickness, and strengthens tight junctions, normalizing gut permeability (“leaky gut”). Mechanisms include endocannabinoid-mediated barrier control and epithelial signaling.
• The Amuc_1100 outer-membrane/pili-like protein enhances transepithelial resistance and signals via TLR2 to elicit IL-10–skewed immune responses, consistent with barrier support and reduced inflammatory tone. Pasteurized cells (which preserve these surface ligands) provide a postbiotic route to barrier tightening—relevant to diet-linked permeability and downstream metabolic inflammation.
• A. muciniphila educates the immune system at baseline: it induces antigen-specific T-cell and IgG1 responses during homeostasis (murine data), a rare property among commensals that aligns with immune “tone” setting rather than overt activation.
• A defined phospholipid from A. muciniphila triggers homeostatic immune responses in vivo, providing molecular proof that specific cell components can shape host immunity.
• Emerging work suggests trained-immunity–like effects after exposure to A. muciniphila, adding a durable layer to immune modulation (preclinical). AKK001’s barrier-first mechanism is complemented by immune calibration—tilting responses toward controlled, IL-10-compatible homeostasis, which is metabolically favorable.
• A. muciniphila secretes a protein (P9) that binds ICAM-2 on L-cells and directly stimulates GLP-1 secretion, improving glucose homeostasis and increasing thermogenesis in mice (↑ UCP1 in brown adipose tissue).
• Pasteurized A. muciniphila also increases energy expenditure and spontaneous activity in mice, consistent with weight-regulatory potential alongside GLP-1 biology.
• GLP-1 signaling is a strong mechanistic candidate (preclinical), while human metabolic wins (insulin/cholesterol) are already demonstrated; weight reduction signals are present as trends and may strengthen under longer duration or in combination programs.
Ansaldo, E., Slayden, L. C., Ching, K. L., Koch, M. A., Wolf, N. A., Plichta, D. R., Brown, E. M., Graham, D. B., Xavier, R. J., Moon, J. J., & Blumberg, R. S. (2019). Akkermansia muciniphila induces intestinal adaptive immune responses during homeostasis. Science, 364(6446), 1172–1177. https://doi.org/10.1126/science.aaw7479
The information presented on this document is for educational and informational purposes only. It is not intended to serve as medical advice, or to replace consultation with a qualified healthcare professional.
The statements made regarding these products and ingredients have not been evaluated by the Food and Drug Administration (FDA). This product is not intended to diagnose, treat, cure, or prevent any disease.
Always consult with your physician or another qualified healthcare provider before making any changes to your diet, supplements, or lifestyle. Individual results may vary.